
第一作者:乔雪姣
通讯作者:余珂
通讯单位:北京大学深圳研究生院
图片摘要

成果简介
近日,北京大学深圳研究生院环境与能源学院余珂团队在环境领域著名学术期刊Water Research上发表了题为“Nitrite impairs bioreactor performance due to decreased replication of Candidatus Brocadia sapporoensis by unbalanced energy allocation”的论文。此研究明确了过量亚硝酸盐对厌氧氨氧化细菌及共生菌的氮素循环、糖类、氨基酸、辅酶和维生素合成强度的影响,阐明了这些影响对厌氧氨氧化体系脱氮性能的控制机制。具体来说,过量亚硝酸盐并非通过抑制厌氧氨氧化途径相关基因表达引发厌氧氨氧化反应体系的崩溃,而是通过破坏Ca. Brocadia sapporoensis的能量合成及能量分配能力,将更少的能量用于细胞保护系统、抑制重要细胞结构(细胞膜及细胞壁)生成及DNA复制相关功能,从而阻碍其细胞复制能力。
引言
厌氧氨氧化工艺以其低碳节能的优势在废水脱氨氮领域展现出了巨大的应用潜力。然而,厌氧氨氧化细菌倍增时间长且对环境因子变化敏感,导致反应器启动缓慢,且受到抑制后恢复困难,这严重限制了其在水环境工程中的广泛应用。亚硝酸盐既是底物又是强抑制剂,是影响厌氧氨氧化过程的最关键因素之一。现有研究报道的亚硝酸盐抑制浓度阈值差异显著(5-750 mg N/L),这种差异源于实验条件的多样性,导致工程实践中难以形成统一的风险防控标准。迄今为止,由于亚硝酸盐的抑制机制尚不清楚,仍然缺乏有效的策略来防止亚硝酸盐引起的反应器崩溃。目前一些研究已经建立了亚硝酸盐冲击的微生物群落演替模式,揭示过量亚硝酸盐通过促进竞争性异养微生物的增殖,降低反应体系的脱氮效能。尽管如此,这一推论主要源于对微生物相对丰度变化的观察,而厌氧氨氧化细菌在物种水平上的代谢变化,特别是与能量代谢、细胞复制等重要功能相关的代谢变化的理解仍然有限。这种限制可能与亚硝酸盐抑制机制密切相关,因为能量代谢是微生物生理学的基石,为广泛的生命活动和物质转化提供动力。且微生物的能量分配容易受到环境胁迫的影响,从而影响其氮代谢和繁殖能力。因此,了解过量亚硝酸盐如何影响厌氧氨氧化细菌的能量分配可能是揭示亚硝酸盐抑制机制的关键。厘清微生物的响应策略可以为制定有效的亚硝酸盐胁迫管理措施,提高废水处理效率和稳定性提供有价值的见解。
图文导读
亚硝酸盐对膜生物反应器性能的影响

图1: (a)进水和出水的氮浓度。(b)氮去除效率、氮去除率和氮负荷率的变化。(c)出水TOC浓度的变化。(d)基于MAGs的门水平群落组成。
在稳定期,出水NO2--N浓度接近0 mg N/L,NO2--N、NH4+-N和TN的去除效率分别在99%、87%和87%以上,反应器脱氮性能稳定。在崩溃期间,出水NO2--N浓度最高升到243 mg N/L(84 mg N/gVSS),氮去除负荷从1900 mg N/L/d以上下降到1400 mg N/L/d左右,TN去除率降至69%,并观察到颗粒污泥解体。使用血清瓶测定厌氧氨氧化体系的脱氮能力后,降低氮负荷并通过缓慢提高的方式对反应器性能进行恢复。
厌氧氨氧化系统脱氮基因表达量及脱氮效率的变化
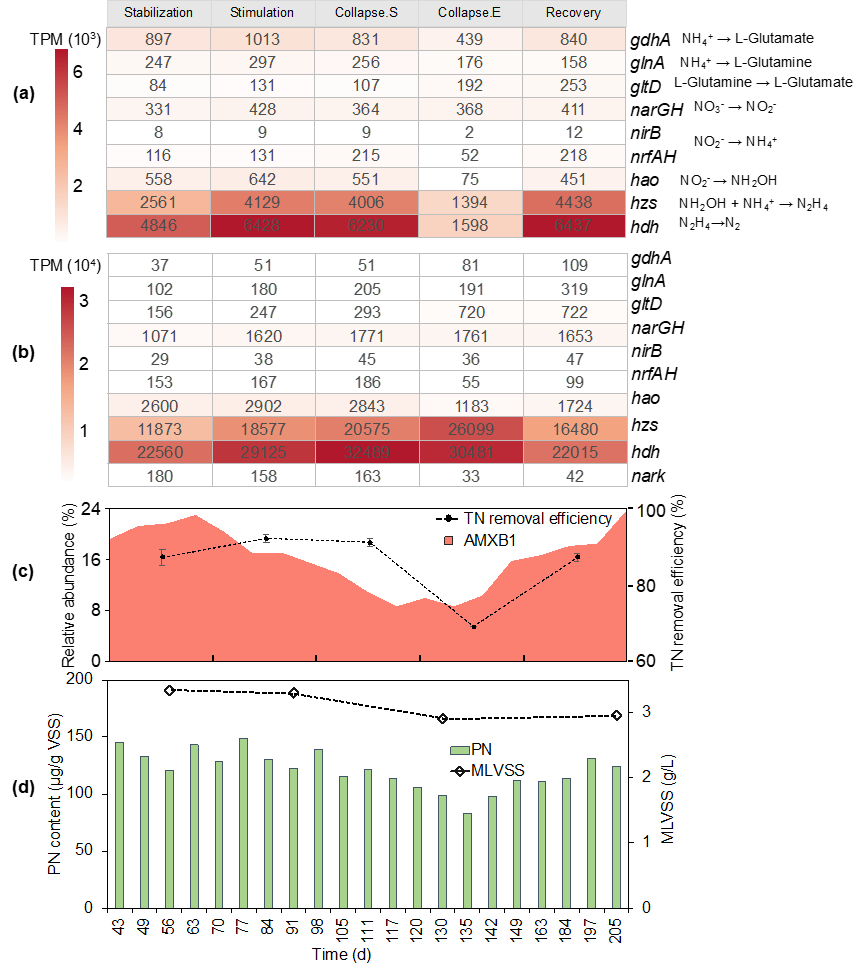
图2:(a)厌氧氨氧化菌群落和(b)AMXB1基因组脱氮基因表达量变化。表达量用TPM表示,为同一期样本的平均值。(c)总氮去除率和AMXB1的相对丰度变化。(d)厌氧氨氧化系统生物量变化。
出乎意料的是,通过计算反应器中厌氧氨氧化细菌Ca. Brocadia sapporoensis (AMXB1)基因组在不同时期的氮代谢基因表达情况(图2b)发现,过量亚硝酸盐并未抑制AMXB1的脱氮基因的表达量,反而能够明显上调anammox途径的基因表达。这可能是AMXB1在面对高浓度亚硝酸盐应激源时,采取的一种对抗或减轻损伤的代偿机制。当考虑了AMXB1生物量(图2d)变化后,对厌氧氨氧化群落进行脱氮基因表达量计算,发现崩溃期间anammox途径的基因表达量明显下调(图2a),这与反应器脱氮性能骤减现象相吻合(图2c)。因此,我们提出假设,亚硝酸盐造成体系崩溃的原因是破坏AMXB1的繁殖能力而非抑制其脱氮基因表达。
亚硝酸盐对AMXB1细胞复制能力的抑制机制

图3: (a)AMXB1在不同时期的基因表达差异概况。(b)AMXB1在不同时期的代谢通路表达的变化倍数(c)AMXB1在不同时期的功能模块表达的变化倍数。其中T1为胁迫期和稳定期的对比,T2为崩溃前期和稳定期的对比,T3为崩溃后期和稳定期的对比,T4为恢复期和崩溃后期的对比。
首先,过量亚硝酸盐破坏了AMXB1的自我保护机制,鞭毛组装、细菌趋向性及双组分系统功能在崩溃期严重下调,导致AMXB1丧失趋利避害能力(图 3b)。其次,构成细胞膜和细胞壁的重要成分如磷脂酰胆碱和磷脂酰乙醇胺、脂多糖等功能被亚硝酸盐严重抑制,导致重要细胞结构合成受损(图 3c)。此外,DNA复制能力被抑制,具体表现为涉及DNA复制的原料ADP、UDP及CDP的合成能力在崩溃期显著下调(图 3c)。然而,更多能量被用于增强氮代谢和碳水化合物代谢包括糖酵解、TCA循环、磷酸戊糖途径(图3c),更少的能量用于鞭毛组装、细胞结构生成及DNA复制。加之在崩溃期间ATP合成减少,更加阻碍了微生物的正常繁殖。因此,过量亚硝酸盐导致的代谢途径能量分配不均阻碍了AMXB1的细胞增殖。
AMXB1及共生菌互养关系的转变
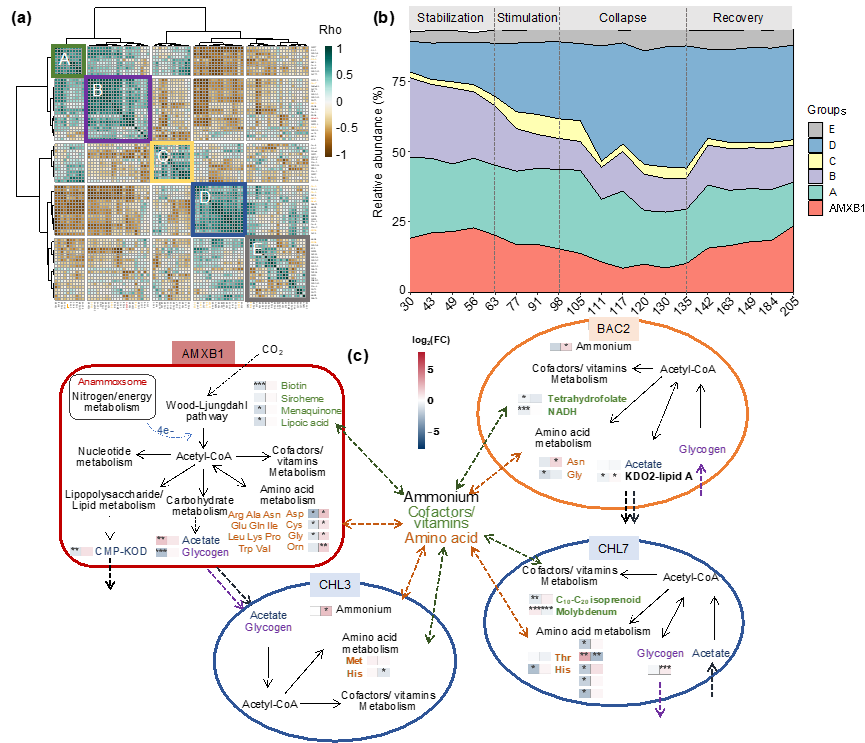
图4:(a)基于微生物相对丰度的聚类热图。(b)不同分组微生物的相对丰度变化。(c)AMXB1及共生菌的互养策略。
厌氧氨氧化细菌并未实现纯培养,共生菌的功能变化也会影响厌氧氨氧化细菌的功能变化。因此,通过对基因组在各个样品中的相对丰度进行聚类(图 4a),B组微生物与AMXB1丰度变化一致(图 4b),说明他们之前存在密切的相互关系,表明AMXB1和B组微生物对反应体系的稳定运行具有至关重要的作用。通过对B组微生物的核心代谢(能量代谢、碳氮代谢、氨基酸合成、辅酶和维生素合成、糖类和脂质合成、物质转运)进行分析,构建了关键物种与AMXB1的互营关系模型(图 4c)。由此发现,过量亚硝酸盐抑制了AMXB1和CHL7的氨基酸(如天门冬氨酸、赖氨酸、亮氨酸和苏氨酸)、辅酶(如钼辅因子、C10-C20 isoprenoid)和维生素(如微生物B6)的交叉喂养。而在恢复期,这些交叉喂养关系逐步恢复,表明这种密切的交叉喂养关系有助于提高必需物质的相互交换,对厌氧氨氧化体系脱氮性能恢复具有重要作用。
小结
本研究解析了厌氧氨氧化菌对过量亚硝酸盐胁迫的分子响应机制(如能量分配、核心代谢转移),并揭示了亚硝酸盐冲击后微生物互养作用在系统功能恢复中的关键角色。主要发现如下:(1)过量的亚硝酸盐会破坏AMXB1的能量代谢,降低用于胁迫耐受性发育的能量。(2)过量亚硝酸盐通过损害DNA复制、细胞膜和细胞壁的形成以及细胞保护系统来破坏AMXB1的细胞复制。(3)过量亚硝酸盐造成反应器崩溃的原因是,阻碍了厌氧氨氧化细菌增殖能力而非抑制其脱氮基因表达。(4)特定的氨基酸和辅助因子,如谷氨酸和生物素,可能会减轻亚硝酸盐的抑制并恢复脱氮性能。
本项目得到了国家自然科学基金委、北京大学AI for Science优先项目、广东省自然科学基金委、深圳市知识创新计划基础研究项目、新西兰皇家学会马斯登基金的支持。
作者简介

第一作者:乔雪姣,北京大学深圳研究生院,博士后,导师为余珂老师。主要从事厌氧氨氧化细菌在复杂废水体系中的脱氮效能强化与运行稳定性提升,重点围绕养殖废水及高盐废水处理场景,在利用多维组学解译厌氧氨氧化菌对抗生素及高盐的适应性机制方面取得了创新性成果。目前,共发表中英文文章21篇,含以第一作者在相关领域Water Research, Environment International, Bioresource Technology等期刊的多篇文章,主持深圳市面上项目1项。

通讯作者:余珂,北京大学深圳研究生院,副教授,环境科学与工程学、计算机应用技术双学科博导。主要从事环境微生物组及生物信息学方法研究,专注于生物信息学分析算法、流程及可视化开发,结合培养组发掘极端环境体系中具有应用潜力的微生物。研究同时关注于多宏组学联用技术开发,解析群落微生物的生态位分异及其互作关系。成果包括论文90余篇,含以第一作者或通讯作者身份在包括Nature Communications, Microbiome, Environmental Science & Technology,Water Research等微生物组学、环境工程等领域的多篇论文。
联系方式:yuke.sz@pku.edu.cn
文章链接:https://doi.org/10.1016/j.watres.2025.123806



